Công ty Cổ phần Thú y Amavac



Tóm tắt:
Dưới dòng 1.5 (sublineage 1.5 hay còn gọi là NADC34-like) của virus gây hội chứng hô hấp và sinh sản ở lợn (PRRSV) mới xuất hiện, đã gây ra mối đe dọa trực tiếp đối với ngành chăn nuôi lợn Trung Quốc kể từ năm 2018. Tuy nhiên, mức độ lưu hành và tác động của chủng NADC34-like đối với các trang trại lợn Trung Quốc chưa được làm sáng tỏ.
Trong nghiên cứu hiện tại, chúng tôi theo dõi các tác nhân gây bệnh bao gồm PRRSV, virus gây bệnh dịch tả lợn châu Phi (ASFV), virus gây bệnh dịch tả lợn cổ điển (CSFV), virus gây bệnh giả dại (PRV) và Porcine circovirus type 2 (PCV2) ở một trang trại lợn vỗ béo, thực hành chăn nuôi an toàn sinh học nghiêm ngặt ở tỉnh Hắc Long Giang, Trung Quốc, từ năm 2020 đến năm 2021.
Kết quả cho thấy nhiều chủng PRRSV cùng tồn tại ở trang trại, trong đó các chủng thuộc nhóm NADC30-like và NADC34-like là những chủng chiếm ưu thế. Đáng chú ý, chủng PRRSV nhóm NADC34-like được phát hiện trong thời kỳ tỷ lệ tử vong cao nhất, là một trong những chủng chiếm ưu thế tại trang trại này.
Kết quả so sánh cho thấy protein NSP2 của các chủng virus có cùng kiểu xóa 100 aa giống như chủng IA/2014/NADC34 được phân lập ở Mỹ năm 2014. Phân tích cây phát sinh chủng loại dựa vào trình tự gen ORF5 cho thấy sự biến đổi không nhiều giữa các chủng NADC34-like lưu hành ở trang trại, với một tốc độ tiến hóa tương đối cao. Phân tích đa hình bằng phương pháp RFLP cho thấy hầu hết kiểu gen ORF5 là 1-7-4, với một mẫu là 1-4-4. Kết quả phân tích trình tự hai bộ gen hoàn chỉnh của PRRSV NADC34-like ở trang trại cho biết hai chủng virus có mức tương đồng nucleotide là 98,9% và không là virus tái tổ hợp.
Tóm lại, đây là nghiên cứu đầu tiên về mức độ phổ biến và đặc điểm của PRRSV nhóm NADC34-like tại một trang trại chăn nuôi quy mô lớn ở miền bắc Trung Quốc. Những kết quả này giúp làm rõ ảnh hưởng của virus nhóm NADC34-like đối với các trang trại lợn ở Trung Quốc và cung cấp tài liệu tham khảo để phát hiện cũng như phòng ngừa đối với virus thuộc nhóm NADC34-like.
1. Đặt vấn đề
Kể từ năm 2018, Trung Quốc_ một trong những chăn nuôi lợn lớn nhất trên thế giới, không chỉ phải đối mặt với virus gây bệnh dịch tả lợn cổ điển (CSFV), virus gây bệnh giả dại (PRV), Porcine circovirus type 2 (PCV2), virus gây hội chứng hô hấp và sinh sản ở lợn (PRRSV), mà còn phải đối mặt với virus gây bệnh dịch tả lợn châu Phi (ASFV). PRRS được xác định là bệnh phổ biến ở các nước chăn nuôi lợn trên thế giới, đặc trưng bởi hiện tượng rối loạn sinh sản ở lợn nái (sảy thai, tăng tỷ lệ thai chết lưu), rối loạn hô hấp ở lợn mọi lứa tuổi, tăng tỷ lệ chết và làm giảm sức sản xuất_ đặc biệt ở lợn cai sữa và lợn con theo mẹ. Cũng do các loại vắc xin tai xanh hiện tại có mức độ bảo hộ khác nhau chống lại đồng chủng và biến chủng, mức độ đa dạng của các chủng virus thực địa dẫn tới rất khó dự đoán về khả năng bảo hộ chống lại các biến chủng, gây ra thiệt hại lớn về kinh tế cho ngành chăn nuôi lợn toàn cầu.
PRRSV thuộc giống Betaarterivirus, họ Arteriviridae, bộ Nidovirales. PRRSV hiện được phân thành hai loài riêng biệt: Betaarterivirus suid 1 (PRRSV-1) và Betaarterivirus suid 2 (PRRSV-2). Cả hai loại PRRSV đã lưu hành ở Trung Quốc trong nhiều thập kỷ và đã gây ra thiệt hại kinh tế đáng kể cho ngành chăn nuôi lợn của Trung Quốc. PRRSV-2 là chủng gây bệnh chính ở Trung Quốc và được phân loại thành 9 dòng dựa trên trình tự gen ORF5. Bốn dòng (1, 3, 5 và 8) của PRRSV-2 được xác định lưu hành ở thực địa. Trong những năm gần đây, các dòng lưu hành chính của PRRSV-2 đã chuyển từ dưới dòng 8.7 (CH-1a-like và HP-PRRSV-like) sang dưới dòng 1.8 (NADC30-like).
Vào năm 2014, dòng ORF5 RFLP 1-7-4 đã phổ biến ở Mỹ và được xác nhận là nguyên nhân gây ra những đợt dịch sảy thai dữ dội ở đàn lợn nái và gây chết cao ở lợn con. Về sau, dưới dòng này cũng được xác nhận lưu hành ở Peru. Ở Trung Quốc, PRRSV thuộc nhóm NADC34-like lần đầu tiên được phát hiện ở tỉnh Liêu Ninh vào năm 2017. Sau đó, các chủng thuộc nhóm này được phát hiện ở miền nam Trung Quốc. Vào năm 2020 và 2021, dưới dòng 1.5 (NADC34-like) tiếp tục được phát hiện và được nghiên cứu về sự lây lan của chúng. Không giống như PRRSV NADC34-like của Mỹ, PRRSV NADC34-like của Trung Quốc hiện đang gây bệnh ở mức độ nhẹ hoặc trung bình. Tuy nhiên, mức độ phổ biến và ảnh hưởng của PRRSV NADC34-like đối với các trang trại lợn ở Trung Quốc vẫn chưa được biết. Trong nghiên cứu này, chúng tôi đã tiến hành theo dõi sự lưu hành của PRRSV trong một chu kỳ sản xuất (150 ngày mỗi chu kỳ) tại một trang trại lợn vỗ béo. Quy mô chăn nuôi của trang trại này là 3400 con. Chúng tôi đã nghiên cứu diễn biến dịch bệnh và đặc điểm phân tử của PRRSV NADC34-like ở trang trại này.
2. Kết quả và thảo luận
2.1. Kết quả phát hiện virus
Tổng cộng có 412/3400 con lợn chết trong vòng 150 ngày theo dõi, với tỷ lệ chết là 12,12% (Phụ lục S1). Tổng cộng có 46 lợn con chết trong 15 ngày đầu tiên, được đặt tên là giai đoạn 1 (Hình 1). Các triệu chứng lâm sàng chính là bỏ ăn và sốt, sau đó là chết cấp tính, và những thay đổi bệnh lý chính được quan sát thấy là xuất huyết ruột, xuất huyết ổ bụng và tràn dịch màng bụng. Nguyên nhân chết này được đánh giá là do nhiễm khuẩn dựa trên các triệu chứng lâm sàng, kết quả xét nghiệm và hiệu quả của can thiệp kháng sinh (Phụ lục S2). Đến ngày thứ 10, kháng sinh được sử dụng khẩn cấp để phòng và điều trị (dựa trên kết quả kiểm tra độ nhạy cho thấy cefoperazone–sulbactam có hiệu quả) đã dẫn tới các triệu chứng thuyên giảm rõ rệt.
Giai đoạn tử vong cao nhất (287/412) xảy ra từ ngày thứ 16 đến ngày thứ 45, được đặt tên là giai đoạn 2 (Hình 1). Hầu hết lợn đều có các triệu chứng hô hấp rõ rệt như ho, thở khò khè hoặc thở thể bụng. Kháng sinh (cefoperazone–sulbactam, doxycycline hydrochloride dạng bột hòa tan, dung dịch tilmicosin và neomycin sulfate) được sử dụng để phòng và điều trị nhưng hiệu quả không khả quan. Mổ khám cho thấy phổi bị đặc, rải rác xuất huyết ở kẽ các tiểu thùy phổi và xuất huyết ổ bụng.
Dựa trên các triệu chứng lâm sàng và kết quả xét nghiệm sau đó, chúng tôi cho rằng những biểu hiện này là do nhiễm virus. Số lợn chết (79/412) giảm sau ngày thứ 45, được đặt tên là giai đoạn 3 (Hình 1a) và phần lớn số chết là lợn đã được cách ly trước đó vì các triệu chứng hô hấp.
Để tìm hiểu nguyên nhân gây chết, tổng cộng 283 mẫu đã được thu thập từ lợn chết và xét nghiệm ASFV, CSFV, PRRSV, PRV và PCV2. PRRSV và PCV2 đã được phát hiện, trong khi ASFV, CSFV và PRV cho kết quả âm tính (Phụ lục S2). Trong ba giai đoạn được ghi nhận ở trên, tỷ lệ phát hiện PRRSV và PCV2 là 17,39% (8/46), 33,13% (53/160) và 29,87% (23/77); và 89,13% (41/46), 81,88% (131/160) và 80,52% (62/77) tương ứng (Hình 1a). Dựa trên các kết quả này, chúng tôi suy đoán nguyên nhân gây chết trong giai đoạn đầu chủ yếu là do nhiễm vi khuẩn, và PRRSV đó đã góp phần gây ra tỷ lệ tử vong cao ở trang trại lợn, còn PCV2 thì không.
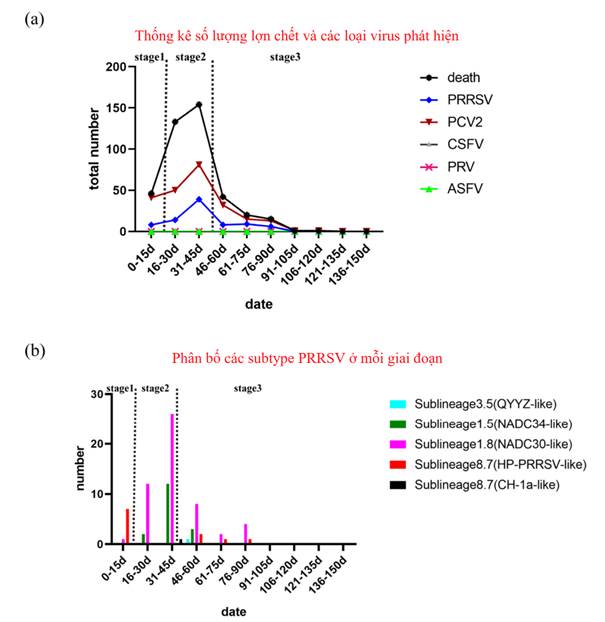
Hình 1. Thống kê số lượng lợn chết và các loại virus phát hiện được
2.2. Phân tích cây phát sinh chủng loại
Để tìm hiểu mối quan hệ giữa các subtype PRRSV và hiện tượng chết của lợn trong trang trại, chúng tôi đã giải trình tự gen NSP2 và ORF5 tất cả các mẫu dương tính với PRRSV bằng phương pháp Sanger.
Tổng cộng có 78 trình tự NSP2 và 69 trình tự ORF5 đã thu được; trong số này, 54 mẫu được xác định là PRRSV NADC30-like (64,29%), 17 mẫu là PRRSV thuộc nhóm NADC34-like (20,24%), 11 là PRRSV thuộc nhóm HP-like (13,10%), 1 là PRRSV nhóm CH-1a-like (1,19%), và 1 là PRRSV thuộc nhóm QYYZ-like (1,19%) (Hình 1b). Do đó, PRRSV nhóm NADC30-like, NADC34-like và HP-like là các chủng chính gây bệnh ở trang trại này.
Hơn nữa, trong 3 giai đoạn dịch, tỷ lệ phát hiện các chủng chính lần lượt là 12,50%, 71,70% và 65,22% (với NADC30-like); 0,00%, 26,42% và 13,04% (với NADC34-like); và 87,50%, 1,89% và 17,39% (với HP-like-PRRSV). Đáng chú ý là thời điểm đỉnh dịch lại trùng với thời điểm phát hiện được sự lưu hành chỉnh của các nhóm NADC30-like và NADC34-like (Hình 1).
Các kết quả trên chứng minh rằng PRRSV nhóm NADC30-like và NADC34-like mà không phải là PRRSV nhóm HP-like (độc lực cao) có liên quan chặt chẽ đến nguyên nhân chết của lợn ở trang trại này. Ngoài ra, một số chủng thuộc nhóm NADC34-like (14/17) vẫn tiếp tục được phát hiện trong 15 ngày sau khi phát hiện ban đầu ở giai đoạn 2. Sự lây lan của PRRSV nhóm NADC34-like dường như nhanh hơn so với PRRSV nhóm NADC30-like. Mức độ độc được báo cáo trước đây ở Trung Quốc là nhẹ và trung bình, và mức độ độc lực của PRRSV nhóm NADC34-like ở các nước trên thế giới cũng rất thay đổi. Khả năng gây bệnh của PRRSV nhóm NADC34-like ở trang trại này vẫn đang được nghiên cứu.
2.3. Phân tích trình tự gen virus thuộc nhóm NADC34-like
Sự xuất hiện của PRRSV nhóm NADC34-like lần đầu tiên được báo cáo ở tỉnh Liêu Ninh, Trung Quốc vào năm 2017. Chủng này sau đó đã xuất hiện ở các tỉnh khác của Trung Quốc. Các mẫu dương tính được giải trình tự thông qua phương pháp Sanger. Tất cả các trình tự NSP2 của các chủng nhóm NADC34-like đều có chung đặc điểm xóa liên tục 100 axit amin ở giữa vị trí 328 và 427 như đã báo cáo trước đây, so với ATCC-VR2332 (GenBank: U87392) (Hình 2). Những sự mất đoạn này có thể được sử dụng làm chỉ thị phân tử để phân biệt các chủng virus thuộc nhóm NADC34-like với các chủng PRRSV type 2 khác ở Trung Quốc, tương tự như kiểu xóa protein NSP2 ổn định ở các chủng PRRSV thuộc nhóm NADC30-like. Mức tương đồng trình tự axit amin protein NSP2 của các chủng virú thuộc nhóm NADC34-like trong trang trại này là từ 99,2% đến 99,9%. So sánh với cơ sở dữ liệu của NCBI, tương đồng cao nhất là với chủng IA/2014/NADC34 (GenBank: MF326985), ở mức 92,6%.

Hình 2. So sánh đặc điểm trình tự amino acid protein NSP2
Mức độ tương đồng nucleotide của gen ORF5 giữa các chủng PRRSV nhóm NADC34-like ở trang trại lợn này là 99,2–100% và cũng tương đồng cao nhất với chủng IA/2014/NADC34 trong cơ sở dữ liệu NCBI (96,9–97,2%). Mức tương đồng giữa các chủng này và chủng virus thuộc nhóm NADC34-like đầu tiên được báo cáo ở Trung Quốc (LNWK130) là 94,9–95,0%. Những kết quả này chỉ ra rằng PRRSV thuộc nhóm NADC34-like ở Trung Quốc đã có sự tiến hóa. Kết hợp với kết quả phân tích trình tự protein NSP2 của PRRSV nhóm NADC34-like, có thể thấy chỉ có 1 chủng virus thuộc nhóm NADC34-like lưu hành ở trang trại này; điều này giúp cho việc nghiên cứu tốc độ tiến hóa của chủng virus thuộc nhóm NADC34-like.
Các chủng virus thuộc nhóm NADC34-like ở trang trại được ước tính có tốc độ đột biến nucleotide là 3,1 × 10−2 đột biến/vị trí/năm—cao hơn tốc độ tiến hóa, nằm trong khoảng từ 6,6 × 10−3 đến 1,3 × 10−2 đột biến/vị trí/năm đối với các subtype thuộc dòng 1 đã được báo cáo trước đây ở Mỹ. Đây là một phát hiện đáng báo động chỉ ra rằng thời gian từ khi xuất hiện PRRSV nhóm NADC34-like ở Trung Quốc đến khi đạt đỉnh điểm trong quần thể đã ngắn hơn so với ở Mỹ (trung bình 4,5 năm). Ngoài ra, việc giám sát PRRSV cho thấy số lượng chủng virus thuộc nhóm NADC34-like rõ ràng đã tăng lên kể từ năm 2020—đặc biệt là vào năm 2021 (dữ liệu chưa được công bố). Do đó, chúng tôi suy đoán rằng PRRSV nhóm NADC34-like đã trở nên phổ biến ở một số vùng của Trung Quốc.
Chúng tôi tiếp tục phân loại các chủng PRRSV nhóm NADC34-like trong trang trại này theo tính đa hình chiều dài đoạn giới hạn (RFLP) của gen ORF5. Mẫu RFLP gen ORF5 của TZJ1277 là 1-4-4, trong khi các mẫu khác là 1-7-4. So với biến thể 1C của dòng PRRSV mới xuất hiện (MW887655) ở Mỹ, TZJ1277 có mối quan hệ chặt chẽ hơn với PRRSV thuộc nhóm NADC34-like đã được báo cáo trước đó (Hình 3a). Phân tích RFLP có những điểm hạn chế, đó là các chủng có mối quan hệ di truyền xa nhau nhưng lại có hiểu RFLP giống nhau, và tính không ổn định của các loại RFLP chỉ trong 10 lần tiếp đời trên động vật. Chính vì sự không ổn định trong đặc điểm này, các nhà nghiên cứu trên thế giới đã đưa ra các cách đặt tên riêng dựa trên tình hình dịch bệnh ở mỗi quốc gia.
2.4. Phân tích trình tự gen bộ gen virus
Giải trình tự toàn bộ bộ gen (WGS) có thể cho biết nhiều thông tin hơn về PRRSV so với phân tích trình tự gen ORF5 dựa vào phương pháp giải trình tự Sanger. Để tìm hiểu mối quan hệ tiến hóa của các chủng PRRSV phân lập với các chủng đại diện, chúng tôi đã giải trình tự hai bộ gen hoàn chỉnh từ chủng virus thuộc nhóm NADC34-like ở trang trại này. Theo nghiên cứu trước đây của chúng tôi, các chủng PRRSV nhóm NADC34-like ở Trung Quốc có thể được chia thành hai nhóm (A và B) dựa trên phân tích cây phát sinh chủng loại toàn bộ bộ gen virus. Phân tích cây phát sinh chủng loại bộ gen cho thấy hai chủng có bộ gen hoàn chỉnh được giải mã (TZJ864 và TZJ921) thuộc về nhóm A (Hình 3b). Tất cả các trình tự thuộc nhóm A đều đến từ tỉnh Hắc Long Giang, Trung Quốc, đây là một trong những khu vực xuất hiện các chủng thuộc nhóm NADC34-like sớm nhất.
Cho đến nay, sáu loại vắc xin PRRS thương mại—CH-1R, JXA1P80, HuN4-F112, GDr180, TJM-F92 và RespPRRS MLV đã được sử dụng rộng rãi ở Trung Quốc. Các vắc-xin sống, giảm độc lực này đều có nguồn gốc từ dòng 5 hoặc dòng 8 và có quan hệ họ hàng xa với các chủng virus thuộc nhóm NADC34-like (dưới dòng 1.5). Lợn được tiêm vắc xin CH-1R MLV một tuần sau khi nhập đàn. Tương đồng nucleotide của TZJ864 và TZJ921 với CH-1R chỉ là 83,3%. Ngoài ra, đã có báo cáo rằng vắc-xin PRRSV giảm độc lực hiện có không mang lại hiệu quả bảo vệ tốt đối với các chủng PRRSV nhóm NADC30-like (dưới dòng 1.8) thuộc dòng 1, NADC34-like (dưới dòng 1.5). Vì vậy, việc phát triển vắc-xin từ virus thuộc dòng 1 là rất cần thiết.
.jpg)
Hình 3. Cây phát sinh chủng loại của PRRSV nhóm NADC34-like: (a) dựa vào trình tự gen ORF5; (b) dựa vào bộ gen virus.
2.5. Phân tích hiện tượng tái tổ hợp
Tái tổ hợp là một cơ chế tiến hóa quan trọng của PRRSV. Cả sự tái tổ hợp giữa các dòng và trong cùng một dòng đã xảy ra, góp phần vào sự xuất hiện của biến chủng mới. Biến thể 1-4-4 của Mỹ dòng 1 có chủng virus bố mẹ là IA14737-2016 (nhóm NADC34-like), và trao đổi các đoạn gen với với các chủng IA/2014/NADC34 và NADC30. PRRSV tái tổ hợp ngày càng được phân lập nhiều kể từ khi PRRSV nhóm NADC30-like xuất hiện ở Trung Quốc. Để tìm hiểu xem chủng PRRSV nhóm NADC34-like phân lập từ trang trại này có phải là virus tái tổ hợp hay không, chúng tôi đã thực hiện phân tích tái tổ hợp trên hai chủng virus là TZJ864 và TZJ921 thu được từ trang trại. Phân tích tái tổ hợp và so sánh trình tự cho thấy chúng không phải là virus tái tổ hợp và có độ tương đồng nucleotide cao (98,9%). Tuy nhiên, sự cùng tồn tại của nhiều dòng PRRSV trong đàn lợn tạo điều kiện thích hợp cho sự tái tổ hợp của PRRSV ở thực địa. Chúng tôi đã phát hiện nhiều chủng PRRSV thuộc nhóm NADC34-like tái tổ hợp từ các trang trại lợn khác từ tháng 6 năm 2021 (dữ liệu chưa được công bố). Các đặc tính và độc lực của virus thuộc nhóm NADC34-like tái tổ hợp này cần được nghiên cứu thêm.
Ở Mỹ đã có báo cáo về thiệt hại kinh tế cao do nhóm NADC34-like tái tổ hợp gây ra. Điều quan trọng là các chủng PRRSV nhóm NADC34-like và NADC30-like tái tổ hợp với các chủng thuộc các subtype khác nhau, dẫn đến độc lực khác nhau/ thay đổi giữa các chủng tái tổ hợp và gây trở ngại lớn trong việc phòng ngừa PRRSV. Trong bối cảnh PRRSV nhóm NADC30-like đã trở thành chủng gây bệnh chính ở Trung Quốc, sự bùng phát của PRRSV nhóm NADC34-like ở các trang trại lợn chắc chắn sẽ dẫn đến sự trao đổi đoạn gen thường xuyên hơn giữa chúng. Vì vậy, chúng ta cần nâng cao nhận thức về tầm quan trọng của việc giám sát liên tục các chủng PRRSV thuộc nhóm NADC34-like, kiểm soát chặt chẽ việc lựa chọn lợn giống và ngăn chặn sự xuất hiện của nhiều subtype PRRSV trong cùng trang trại.